
自然林更新主如果指植物从着花, 安谧到种子的分娩、扩散和萌生ai换脸 刘亦菲, 以及幼苗的假寓、存活、孕育和建成的生态学历程[1-2]。其中, 幼苗被合计是丛林更新历程对外界环境最敏锐的时期[3-4], 其建植胜仗与否顺利关系到改日植物群落的更新[5]。与此同期幼苗假寓对植被的更新具有筛选作用[6], 通过影响植物种群的数目构成、漫步模式来影响森群落动态变化, 最终决定了丛林的长期演替模式, 维系了丛林生态系统植被的各种性[7]。
空间模式与生态学历程之间的互相作用是群削发落发展的能源[8]。幼苗四肢林下生态系统的垂死组分, 是木本植物糊口史中死活历程和动态特色最昭彰的时期[9], 其空间漫步模式会对幼苗的自然更新历程会产生垂死的影响。在此历程中, 一些机制大致促进生物各种性的守护[5]。Swamy等合计, 把柄Janzen-Connell假说, 树种的更新模式老是偏畸并远隔于母树[10]。这是由于寄主特异性天敌, 如病原体、专性植食者等, 容易从成年个体扩散到离它们较近的后代; 同期, 由于大多数树种的种子散布在母树周围, 漫步于母树隔壁的同种幼苗个体过于密集, 病原体和专性植食者的繁衍很可能会加重, 使得幼苗因负密度制约(Negative density dependence)而导致多数物化[11-12]。这两个历程使幼苗的存活率随离母树或同种成树距离的增多而升高, 在制约同种幼苗增补的同期, 为其他物种的幼苗更新提供了空间, 从而守护了物种共存。此外, Queenborough等也合计幼苗的空间模式对物种的长期漫步模式有着垂死影响[13]。
另一方面, 丛林的更新模式与林分的空间异质性有垂死的关系[8]。在不同类型的群落中, 地上成体植株的空间异质性为幼苗的增补、定植等历程提供了垂死基础, 影响着系数这个词群落的更新特征和演替空间模式[14]。但是, 以往对幼苗空间漫步模式的探究多集结于特定群落类型[15-17], 对不同演替阶段群落中幼苗空间漫步模式的探究鲜见报说念。
以阔叶红松(Pinus koraiensis)林为演替顶级的次生针阔混交林是我国北温带地区垂死的地带性植被类型, 吉林省蛟河地区是阔叶红松林的主要漫步区之一。早期的东说念主为干预, 如过渡采伐、放牧等步履使多数原始丛林遭到龙套, 退化成次生林群落。比年来在国度“自然林保护”与“退耕还林”等生态战略的推进下, 这些林分形成了针阔混交林次生演替历程中的不同群落类型。尽管如斯, 丛林的自然更新仍然是当地植被复原历程中靠近的重浩劫题。因此, 本文以吉林省蛟河地区不同演替阶段的针阔混交林固定监测样地为基础, 对2012-2014年蚁合3 a的木本植物幼苗拜访数进行了初分析, 旨在:(1) 探讨木本植物幼苗的空间漫步法律阐扬偏激年际间互异; (2) 探讨木本植物幼苗和成树在空间漫步上的相干性法律阐扬。以揭示温带地区不同演替阶段丛林群落的更新法律阐扬, 为该地区生态复原提供表面基础。
1 探究地概况与探究措施 1.1 探究区概况蛟河林业推行区贬责局林场位于吉林省蛟河市前进乡境内(43°51′-44°05′N, 127°35′-127°51′E, 海拔330-1176 m), 属长白山系张广才岭山脉。该地区植被属于长白山植物区系, 自然次生针阔混交林和原始阔叶红松林是当地的主要植被类型[18]。乔木层树种主要包括红松、杉松、东北枫、五角枫、胡桃楸(Juglans mandshurica)、水曲柳和紫椴(Tilia amurensis)等。林下灌木层树种主要有暴马丁香(Syringa reticulatavar.mandshurica)、毛榛(Corylus mandshurica)、东北山梅花(Philadelphus schrenkii)和刺五加(Acanthopanax senticosus)等。主要草本有好意思汉草(Meehania fargesii)、狭叶荨麻(Urtica angustifolia)、荷青花(Hylomecon japonica)、华东蹄盖蕨(Athyrium niponicum)和猴腿蹄盖蕨(Athyrium brevifrons)等。该地区处于中纬度北温带, 属受季风影响的温带大陆性形状, 春季干旱而多大风, 夏令燥热而多雨水, 秋季褊狭而多好天, 冬季清冷漫长。年平均气温3.8℃, 最热月7月日均温为21.7℃, 最冷月1月份平均温度-18.6℃。降水多集结在夏令, 约为700-800 mm。泥土类型主要为暗棕色丛林土[19]。
1.2 样地竖立与拜访把柄CTFS (Center for Tropical Forest Science)的期间表率(), 2010年在探究区内把柄林分的发育阶段和树种构成, 选取3种不同发育阶段的针阔混交林竖立固定监测样地, 包括中龄林(21.84 hm2, 420 m×520 m)、老到林(42 hm2, 500 m×840 m)和老龄林(30 hm2, 500 m×600 m), 样地概况如表 1所示。3块样地形成了一个由次生林迟缓向顶级植被类型过渡的典型演替系列。拜访记载各样地内系数胸径(DBH)≥1 cm的植株个体物种称呼、胸径(cm)、树高(m)、枝下高(m)和冠幅(m), 并给每个植株挂牌、定位。
1.3 幼苗样方竖立2011年7月遴选机械布点的措施, 在3个样地内共竖立了451个“种子雨-幼苗”不雅测样站, 其中中龄林99个、老到林209个、老龄林143个(图 1)。每个样站由1个0.8 m×0.8 m的种子雨收罗器偏激四周距其1 m处的4个1 m×1 m幼苗小样方构成(图 2)。
1.4 幼苗拜访对苗区内胸径小于1 cm的木本植物幼苗进行拜访, 记载每棵幼苗的物种称呼、年事、高度和基径, 同期对每颗幼苗进行挂牌秀气。幼苗的年事是把柄芽鳞痕的个数来笃定。幼苗高度为自然气象下基部到最高顶芽的距离。2012年7-8月进行了第1次幼苗拜访责任, 以后每年7-8月复查拜访1次, 并记载幼苗的出身和物化气象[20]。本文选取2012-2014年间的3次幼苗拜访数据进行分析。
1.5 数据分析措施把柄各样站内的幼苗数目(N)离别为不同的等第: N0=0; 0 < N1≤5; 6≤N2≤10; 11≤N3≤15; 16≤N4≤20; 21≤N5≤25; 26≤N6≤30; 31≤N7≤35; 36≤N8≤40; 41≤N9≤45; 46≤N10≤50; 51≤N11≤55; 56≤N12≤60; 61≤N13≤65; 66≤N14≤70; 71≤N15≤75; 76≤N16≤80; 81≤N17≤85; 86≤N18≤90; 91≤N19≤95; 96≤N20≤100; N21 > 100ai换脸 刘亦菲。统计各样站内每个等第的幼苗个体数。
选取Syrjala测验[21], 来分析样站法式幼苗密度和丰富度的空间漫步在年际间的互异。Syrjala测验是通过贪图两个样本的Cramer-von Mises测验来分析两样本之间的空间漫步互异。Syrjala测验的零假定是在系数这个词探究区域内, 两样本的正态漫步是一致的。设在取样点k的坐标为(xk, yk), di(xk, yk)为第i个种群k点的密度值。构建一个零丁于种群大小的测验, 所不雅测到的密度数据最初被法式化:
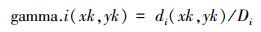
式中, Di是di(xk, yk)不雅测值的总和。第i个种群在点(xk, yk)处的积贮漫步的值, 也即是GAMMA.i(xk, yk), 是系数正态密度不雅察值的总和, gamma.i(xk, yk)。由Syrjala测验零假定的统计量为两个种群之间积贮漫步的差值的平素, 然后系数样本点乞降, 即,
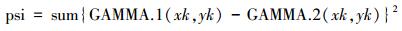
psi是把柄探究区域矩形的角点而转变的, psi被贪图4次, 每1个角点1次, 所得的平均psi值用来作念测验提供假。不雅察到的psi显耀性法式是由它在系数2K个成对枚举所得的测验统计值中的序诸君置而决定的。
本文中所罕有据分析与绘画均遴选海外通用软件R-3.2.0 (-project.org)和Microsoft Excel 2010完成。
2 成果与分析 2.1 种子雨构成2012-2014年间, 中龄林共收罗到14个物种的种子, 从属于8科10属; 老到林共收罗到21个物种的种子, 从属于11科15属; 老龄林共收罗到20个物种的种子, 从属于11科14属20拔擢物(表 2)。3个样地中, 多数物种的种子雨密度皆是在2013年达到峰值, 但也有少数物种如中龄林中的杉松、胡桃楸, 老到林中的辽椴和老龄林中的令嫒榆等的种子雨密度却在2013年最低(表 2)。
2.2 幼苗构成的空间漫步探究期间, 3个样地内拜访到的木本植物幼苗均为该样地存在的主要树种, 莫得发现其他树种的幼苗。样地内幼苗总额皆在2013年达到最大, 2014年幼苗数目最少, 约为2012年、2013年幼苗数目的一半(表 3)。2012-2014年间, 中龄林共监测到10科11属18种木本植物幼苗, 其中2012年拜访到16种1338株幼苗, 莫得发现辽椴(Tilia mandshurica)和令嫒榆; 2013年拜访到18个物种的幼苗, 共1942株; 而2014年共拜访到724株17种幼苗, 莫得发现辽椴(表 3)。老到林共监测到9科11属17种木本植物幼苗, 其中2012年拜访到17种幼苗, 共4041株; 2013和2014年分别拜访到5160和2379株幼苗, 但皆莫得发现毛榛(Corylus mandshurica, 表 3)。老龄林3年内均拜访到16个疏导树种的幼苗, 从属于9科10属, 幼苗株树分别为2199、2229和917(表 3)。
各样地树种幼苗数目与出现的样方数相比成果标明:水曲柳、五角枫和东北枫等幼苗数目较多的物种, 出现的样方数也相对较多(表 3)。3次拜访期间, 中龄林水曲柳幼苗数目最多, 出现样站的比例均在61.6%以上, 其中2012年和2013年出现的样站数昭彰多于2014年; 老到林水曲柳、五角枫和东北枫出现的样站数分别达到总样站数的59.8%、72.7%和37.8%以上; 老龄林中五角枫和东北枫出现的样站数分别达到总样站数的62.2%和48.3%以上。3个样地中, 尽管2012年的杉松幼苗数目远高于五角枫, 但其出现的样站数昭彰小于五角枫(表 3)。
把柄幼苗出现样站相对频率漫步可知:在2012-2014年间, 3个样地均分绝大多数样站出现幼苗数为1-10株, 况兼皆呈现出偏峰场合; 2014年3个样地均以幼苗数为1-5株的样站最多, 况兼幼苗数为0株和1-5株的样站数目均昭彰高于2012年和2013年; 老到林在2013年幼苗数为6-10株的样站数最多, 而老龄林在2012年幼苗数为6-10株的样站数最多(图 3)。
对样站中出现幼苗的物种数进行统计发现, 中龄林在3次拜访期间物种的频率漫步均出现偏锋场合, 而且年际间互异昭彰: 2012年有6个样站中莫得发现任何树种幼苗, 85个样站(占样站总额的85.9%)中出现的物种数小于5; 2013年出现1-5个幼苗物种的样站数最多, 为29个, 远高于2012年和2014年拜访成果; 2014年有18个样站中莫得出现任何幼苗, 61个样站(样站总额的61.6%)发现0-2个物种(图 3)。而老到林和老龄林物种频率漫步的年际互异不昭彰, 且均为正态漫步, 样站数最多的为3-4个树种(图 3)。
2.3 幼苗空间漫步互异遴选Syrjala测验分析了吞并样地内幼苗个体总额、物种数以及4个主要物种幼苗多度的年际间空间漫步互异(表 4)。成果标明:中龄林幼苗物种数和个体数的空间漫步在2013年和2014年存在显耀互异; 水曲柳、杉松和东北枫幼苗数目空间漫步在年际间存在显耀互异, 而五角枫幼苗数目在年际间互异不显耀, 标明五角枫幼苗的产生和漫步在中龄林中相比安谧。老到林幼苗物种数和幼苗个体总额的空间漫步存在显耀的年际互异, 水曲柳、五角枫和东北枫幼苗数目的空间漫步在年际间互异显耀, 而杉松幼苗数目在年际间互异不显耀, 标明杉松幼苗的产生和漫步在老到林中相比安谧。老龄林中, 2012年幼苗个体总额的空间漫步与2013年、2014年互异显耀; 2012年杉松幼苗数目空间漫步与2013年互异显耀; 而幼苗的物种数以及水曲柳、五角枫和东北枫幼苗数目的空间漫步在年际间变化不显耀, 标明老龄林林下幼苗的空间漫步比中龄林和老到林相对安谧。
2.4 主要树种物化幼苗与成树空间漫步对3个样地内水曲柳、五角枫、杉松和东北枫的物化幼苗和成树空间漫步进行相比不错发现:尽管3个样地中物化的幼苗基本漫步在母树隔壁, 况兼整躯壳局与母树保合手一致, 但物化幼苗的空间漫步模式在年际间存在一定互异(图 4-图 6); 2013-2014年间五角枫和东北枫的物化幼苗在3个样地中漫步较为庸碌(图 4, 图 5); 老龄林中, 杉松在2013-2014年间物化的幼苗在样地中漫步的较少(图 6)。
3 议论 3.1 幼苗的数目漫步群落幼苗与地上成体植株的空间漫步模式决定了群落更新特征, 成体植株的空间漫步特征对幼苗建成具有垂死影响[14]。因此, 幼苗的空间漫步对群落更新具有垂死酷爱。本文通过对蛟河地区不同演替阶段针阔混交林样地内木本植物及幼苗的拜访分析, 发现3个样地内拜访到的木本植物幼苗均为该样地存在的主要树种, 莫得发现其他树种的幼苗。木本植物幼苗的数目漫步在不同演替阶段之间皆阐扬出昭彰的互异:水曲柳在演替初期和中期的林分中出现频率较大, 五角枫在演替中期和后期的林分中中出现频率最大, 东北枫在3个样地中出现频率皆相比大; 而胡桃楸、杉松和红松的幼苗在3个样地中出现的频率皆很低。这一方面可能是由于不同演替阶段的林分中, 树种的数目和构成存在互异, 使得不同物种的幼苗漫步在不同群落间存在互异; 另一方面, 啮齿类动物和东说念主类对红松、胡桃楸等种子的取食和采摘也会对幼苗的漫步形成不同进度的影响。探究期间在3个样地内水曲柳、五角枫和东北枫的种子数目在系数树种中均排在前5位, 而胡桃楸、杉松和红松的种子数目较少(未发表数据), 这与陈贝贝等部分探究成果一致[22]; 此外, 幼苗的空间漫步可能与种子的传播特色相干, 水曲柳、五角枫和东北枫的种子主要通过风力传播, 精采的扩散才略也能使得种子和幼苗在样地内庸碌漫步[23], 而胡桃楸、红松和杉松的种子主要通过重力传播, 扩散才略有限。
汤唯车震 3.2 幼苗密度与丰富度的空间异质性本探究成果显现, 在4 m2样站法式上3个不同演替阶段的群落中幼苗密度存在着较大的空间变异性, 而幼苗丰富度的空间异质性较低。这与李晓亮等对西双版纳热带丛林树种幼苗空间漫步探究的部分红果一致[24], 他们合计幼苗密度的空间变异是由微生境的空间异质性导致的, 微生境的空间变异又会反来影响幼苗的存活[25]。而物种数目由于受群落物种区系限定, 空间异质性较低[14]。也有学者合计丛林中幼苗空间模式可能比其周围的环境身分更垂死[16], 因为幼苗密度的空间模式会影响到幼苗的存活气象[24]。举例, Janzen和Connell[11-12]合计漫步于母树隔壁的幼苗由于受到负密度制约效应会阐扬出较高的物化率, 而本探究中3个样地内主要树种的幼苗皆集结于母树隔壁(图 3-图 5)。同期, 以往的探究仍是阐述了负密度制约效应是该探究地区幼苗存活的主要启动机制[20], 这在一定进度上不错反应出幼苗漫步的空间异质性对幼苗密度的影响。
3.3 幼苗空间漫步的年际互异不同庚份之间幼苗个体数和物种数的空间漫步是存在互异的。这主要与种子的产量与扩散相干。最初, 种子是幼苗更新的开首, 幼苗的数目和漫步很猛进度上依赖于种子的可得到性[26], 种子数目多的物种跟容易扩散到更多的样站[27]。另一方面, 不同物种的种子扩散方式有很大互异, 有些物种的种子在阅历了种子雨扩散之后, 还会阅历二次扩散。二次扩散不错镌汰种子的集合进度, 使种子距离母树更远。因此, 种子扩散会导致种子萌生位置的转变进而影响幼苗的漫步。同期, 种子具有就寝的特色, 只须光照、温度等环境身分相宜的时分种子才会萌生, 这也影响到幼苗的漫步模式。除此除外, 不同演替阶段群落中的物种构成和林分结构及环境皆有很大的互异, 因此各林天职幼苗空间漫步在年际间的互异也不尽疏导。
3.4 物化幼苗空间漫步与成树关系除了种子产量和种子扩散方式, 成体植株的空间漫步特征也大致影响幼苗的空间漫步模式[28]。本探究中, 在不同演替阶段的群落内, 水曲柳、五角枫、杉松和东北枫这4个主要树种的物化幼苗在不同庚际间的空间漫步模式与其母树漫步基本一致。这可能是由于选取的4个物种的母树在群落中有较多的个体, 产生的种子量较多, 使母树隔壁有饱和的种源, 进而使得幼苗与母树有着相同的空间漫步模式[24]; 另一方面, 由于种子的传播限定, 大多数树种并不行把种子散布到离母树较远的场合[29-30], 相背, 跟着与母树距离的增多, 种子密度会迟缓镌汰[31], 自然这4个树种的种子皆依靠风力传播, 但是传播的距离并不远, 促使幼苗集合在母树周围。因此, 在负密度制约效应的作用下, 临近母树的幼苗更容易物化。此外, 五角枫、东北枫和沙松的物化幼苗的空间漫步在年际间存在互异ai换脸 刘亦菲, 这可能与幼苗的密度相干。3个样地中, 五角枫和东北枫在2013-2014年间物化幼苗的空间漫步要昭彰大于2012-2013年, 而这两个物种在2013年的幼苗密度均大于2012年, 这从侧面进一步阐述了负密度制约效支吾幼苗空间漫步的影响。